公共サイン納品実績全国3.5万件・植物名ラベル納品実績全国500万枚。Abocは『環境サイン®』・樹名板のトップブランドです

- ホーム
- ネイチャーコンテンツ
- Aboc Museum
- 花の美術館
- 第19回 『黒船が持ち帰った植物たち』
- 1. 黒船による日本及びその付近での植物採集
- 2. 東アジアと北アメリカの間の植物の隔離分布について
- Disjunct distribution of flowering plants...
Disjunct distribution of flowering plants...
Disjunct distribution of flowering plants between eastern Asia and North America (A summary account)
As early as 1846 Asa Gray, then professor of botany at the Harvard University, first noticed the striking and peculiar resemblance between certain plant species of eastern North America and those of Japan, discontinuously occurring some 12,000 km apart. In the course of identifying good series of Japanese plant specimens that were gathered by the U. S. North Pacific Exploring Expeditions he was able to carry out detailed and extensive floristic comparison between Japan and temperate North America, whereby he enumerated more than 580 species, representing over 40 genera, of flowering plants that exhibited the disjunct floristic link between the two remote regions of the world. Since then a number of systematic botanists and phytogeographers, including Fernald, Hultén, Li, Hara, Koyama and Kawano, have pursued investigations on this intriguing phenomenon, some employing palaeobotanical or cytogenetical approaches to supplement basic comparative morphological methods.
At present this particular group of plants is called the Arctic Tertiary Floral Elements, and their phytogeography is generally interpreted as mentioned below.
1. Four types of distributional patterns.
The following are the four geographic types of distributional ranges exhibited by the components of the Eastern Asiatic and temperate North American disjunct flora.
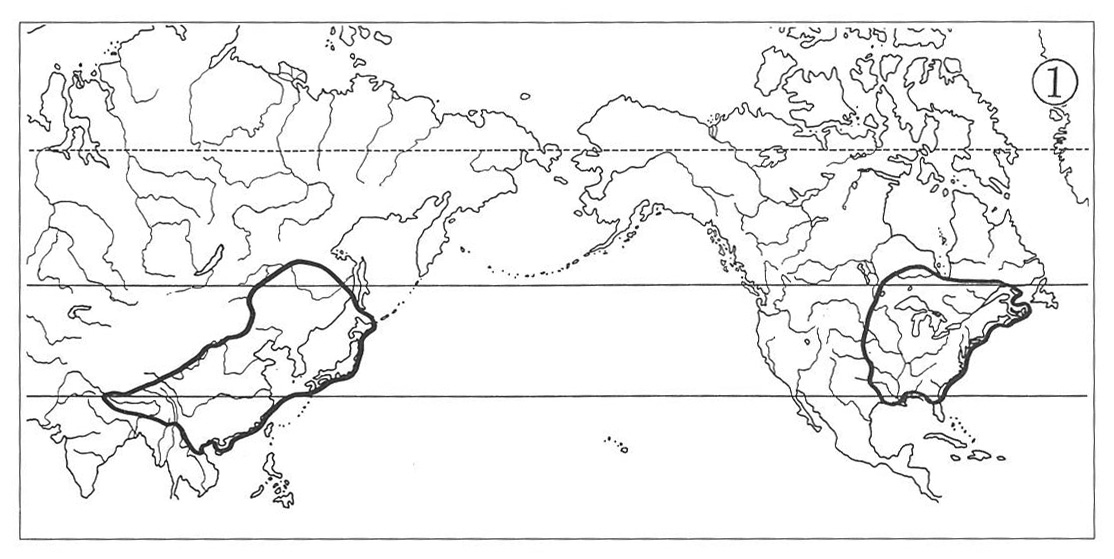
(1) Eastern Asiatic-eastern North American Type:
The North American ranges in this type of distribution encompass the entire Appalachian Mountain Range, frequently extending northwestward to include the Great Lakes Region as well as westward into the eastern part of the Mississippi Basin, while the Asiatic ranges include the whole Japanese Archipelago (from Hokkaido to Kyushu), Amurland, northeastern and central China occasionally with its western extension reaching eastern Himalaya.
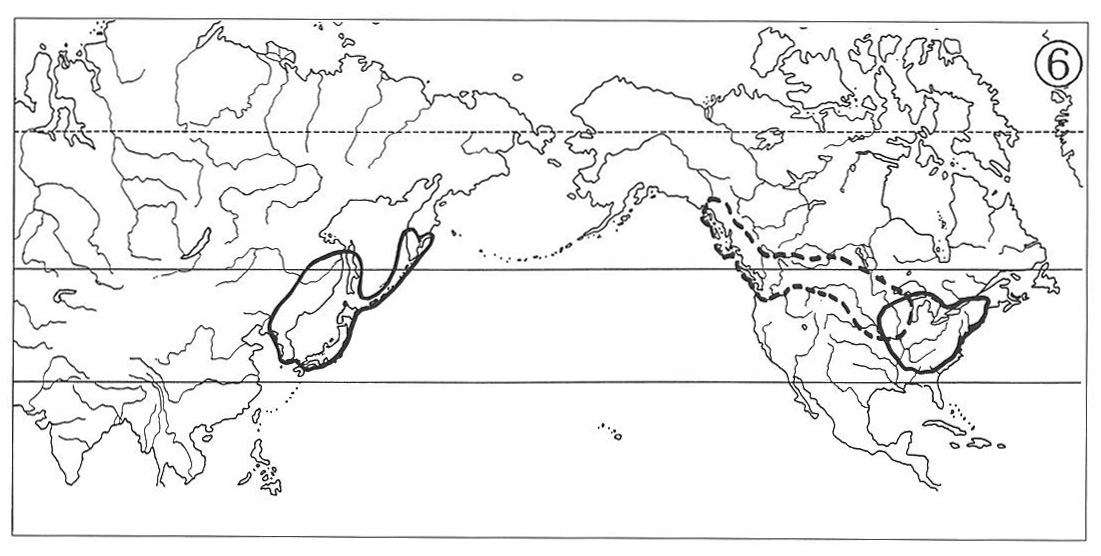
Good examples of plants showing this pattern of distribution include the genera Panax (Map 1), Symplocarpus, Zizania, Hamamelis, Wisteria, Pachysandra, Caulophyllum and Phryma, which are seen in association with either temperate forests or in marshy habitats.
Such genera as Magnolia, Lindera, Croomia and Disporum (Map 2) also belong here, but their ranges of distribution lie more southerly entering warm-temperate to subtropical regions.
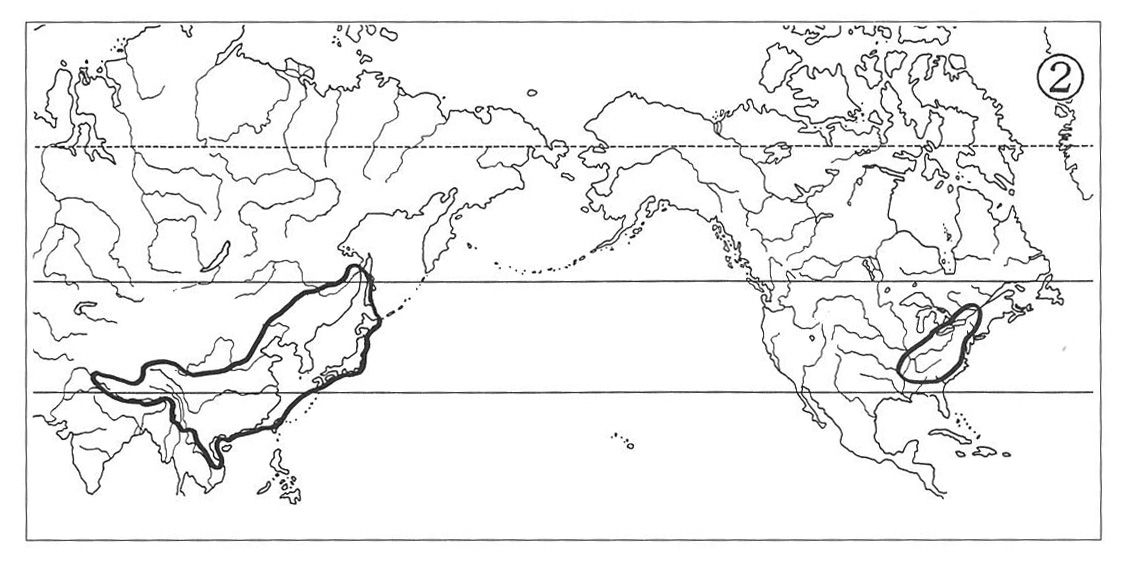
(2) Eastern Asiatic-Western North American Type:
The North American ranges of this type of distribution are situated along the Pacific Coast, covering the Coast Mountain Ranges of British Columbia and its adjoining southern Alaska, Alexander Archipelago, Queen Charlotte Islands and Vancouver Is.
With their southern portions usually expanding to include greater parts of Washington and Oregon as well as northern California, while their northern tip sometimes stretching into Alaskan Peninsula. The Asiatic ranges are, however, essentially the same as those of the type (1). Beside the subgenus Metacoptis of the genus Coptis (Map 4) the genera Lysichiton (Map 3), Adenocaulon, Achlys and Oplopanax exhibit similar ranges of distribution, whereas the range of the genus Castanopsis is located more southerly in warm-temperate regions.
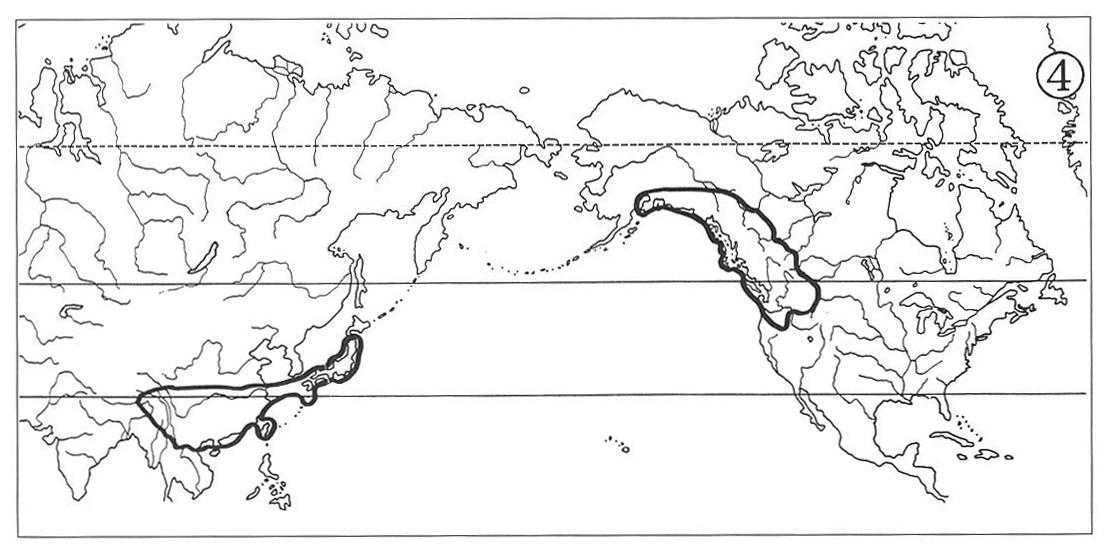
(3) Eastern Asiatic—Eastern and western North American Type:
This type of distribution appears as a combination of the types (1) and (2) above, i.e., the North American ranges develop in both the eastern and the western parts of temperate North America, widely separated by the central and western portions of Mississippi River Basin and continued Rocky Mountains. The counterpart ranges in eastern Asia are similar to those of the two previous types. The genus Clintonia shows a typical example of this distributional pattern (Map 5). The distribution of Arabis lyrate (Map 6) may be attributed to this type. In this species, however, the ranges of its eastern American and western American subspecies meet in western Great Lakes Region, hence forming a continuous specific range simulant of the type (4) below.
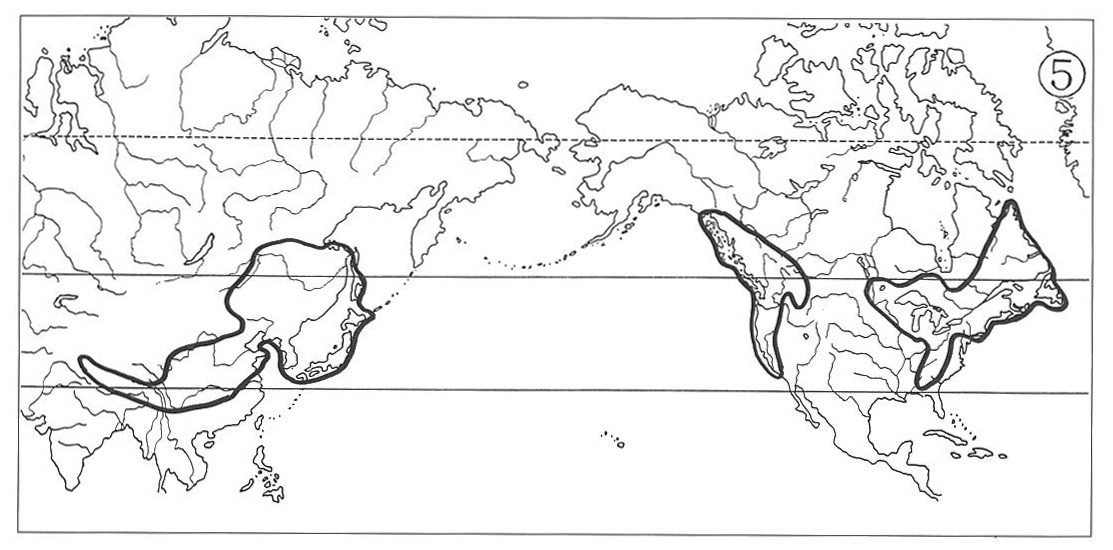
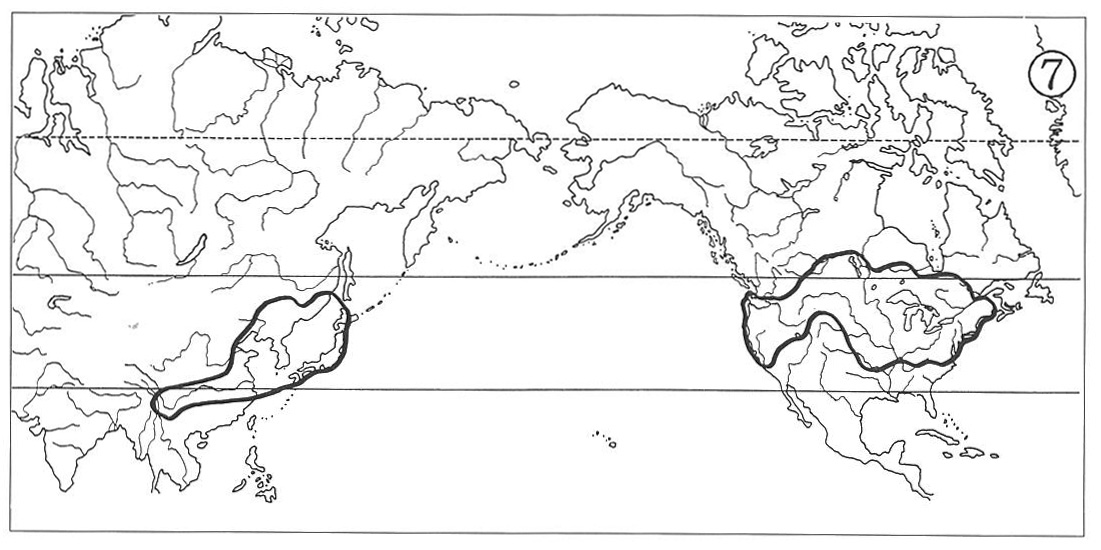
(4) Eastern Asiatic-Trans-North American Type:
In this distributional type the North American ranges extend across southern Canada and adjacent United States northwestwards to Yukon and occasionally as far as to southeastern Alaska. They fill mainly in the glaciated area, and consequently this indicates that after the final withdrawal of the Wisconsin Ice Sheet, plants migrated from temperate United States northwards into northern Quebec and to the Mackenzie District in some instances. The Asiatic counterpart ranges are also relatively larger than those of the previous three types, suggesting that the plant migration here took place from Japan and perhaps Amurland northwestwards to eastern Siberia and/or westwards to Chinese subcontinent.
We encounter those species demonstrating such wide distribution in certain species of Cyperaceae and Gramineae. For instance Bolboschoenus fluviatilis (Cyperaceae) (Map 7) and Beckmannia syzigachne (Gramineae) of marshy habitats, and Carex senanensis (Cyperaceae) and Schizachne purpurascens (Gramineae) in association with northern needle-leaved forests, all suggesting successful ecological adaptation to achieve post-glacial rapid migration.
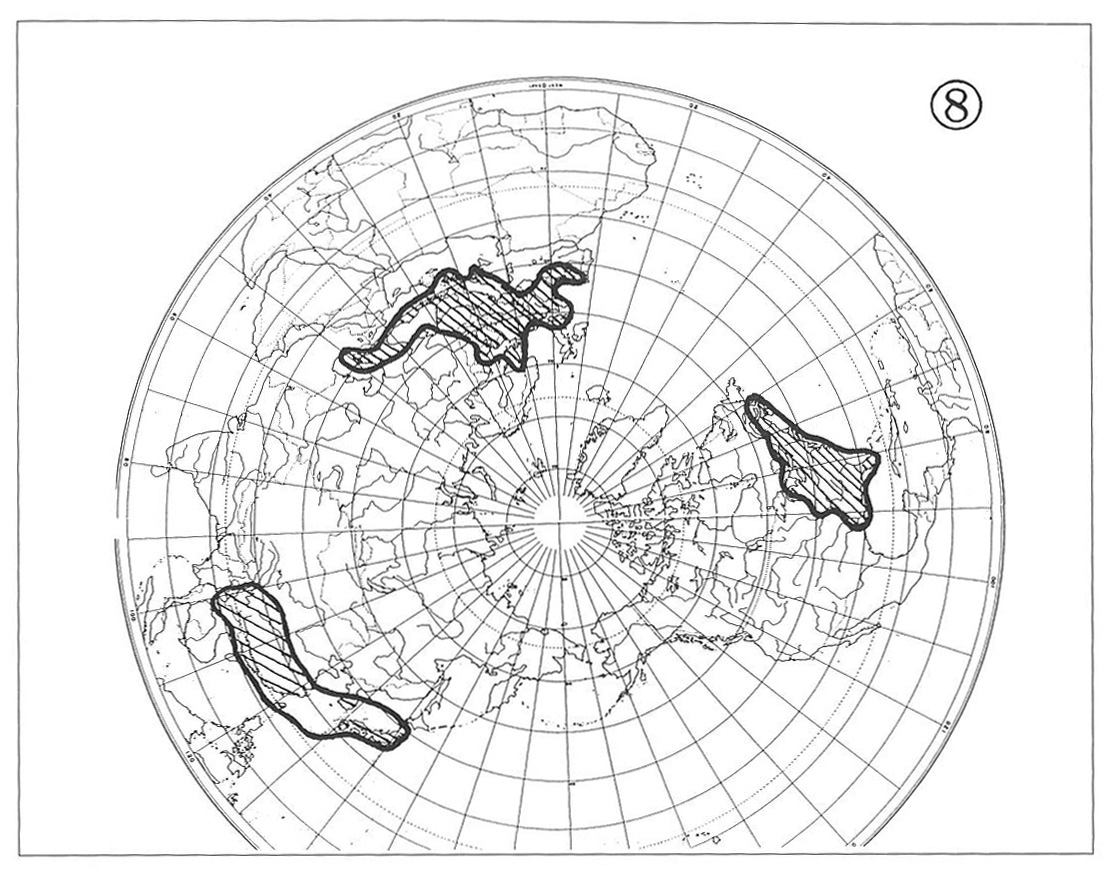
Furthermore, certain components of the Arctic Tertiary Flora are assumed to have developed the so-called circum-polar/boreal ranges of distribution, which after glaciations would have resulted in the discontinuous distribution ranges among Europe, Asia and North America as exemplified by the genera Fagus (Map 8) and Erithronium.
2. Outlines of the history of Arctic Tertiary floral elements.
(1) Origin of the floral elements/and the discontinuous ranges of distribution.
As the name, Arctic Tertiary Flora, signifies, it is believed that the plants now exhibiting the discontinuous distribution between eastern Asia and temperate North America are descendant from the components of the Tertiary flora of the arctic to subarctic region of what are now eastern Siberia, Alaska and their adjacent territories.
This theory is well substantiated by the fossil data obtained from the Tertiary layers, according to which the whole vast territory embracing the middle zone of Siberia, northern China, Sakhalin, as well as in North America, Alaska and the adjacent territory, was in the first half of the Tertiary period, at least as late as the Lower Miocene, under the domination of a summer-green forest flora represented by such genera as Fagus, Ulmus, Alnus, Corylus, Populus, Juglans, etc. (Kryshtofovich 1929).
Therefore, it is assumed that the temperate forest flora had a more or less continuous range over the Beringean district during certain periods of the Tertiary when the Eurasian continent was connected with North America by the Alaska-Beringean landbridge.
Entering the Glacial Epochs, as severe cold climate began prevailing over the Northern Hemisphere, the Tertiary temperate flora would have migrated southwards and the distributional ranges of many of its components would have been separated into Asian and American portions by the Pacific Ocean, hence to form disjunct ranges on the two continents. During the repeated glacial and interglacial stages they would have repeated southward and northward migrations, in the course of which some of them might have differentiated into new taxa, which would have been similar to but slightly different from the original taxa, while some others would have become extinct. Under the severest glaciations such as Würm (Wisconsin), Riss (Illinoian), Mindel (Kansan) and Günz (Nebraskan) they would have survived only in very limited areas along the lims of the continents and in Japan etc., the areas that were not glaciated, i.e., ‟refugia.” The contemporary areas are the results of their expansion of distribution during the post-glacial period.
(2) Ecology of Arctic Tertiary floral elements.
Greater and essential components of the Arctic Tertiary floral elements consist of broad-leaved deciduous trees, forming temperate summer-green forests, and those herbaceous plants growing in association with such deciduous forest vegetations. Minor components, however, include plants of swampy habitats such as Symplocarpus, Zizania or Bolboschoenus, as well as those like Coptis and Schizachne which occur in upper temperate zone and in subarctic needle-leaved forests. The speed of post-glacial migration/expansion of temperate forest species are, as a rule, much slower than in those growing in swampy habitats or in subarctic needle-leaved forests, demonstrating an interesting fact that the mode of distribution of plants has close relevance to habitat factors and vegetation types.
An interesting characteristic of temperate deciduous forests is the existence of conspicuous climatic seasons. In the spring, when the full sunshine reaches the forestfloors through defoliated woods, herbaceous plants of the forestfloors, forming the so-called spring flora, blossom almost simultaneously. Among them are Trillium, Polygonatum, Erythronium, Panax, Viola, Corydalis, Primula and Symplocarpus, for instance. Majority of them are geophytes, and their upperground parts complete seed production and other metabolic activities by early summer and disappear from the ground at that time, when the tree crowns bearing fully developed leaves intercept the strong summer sunshine from the forest floors. In the summer, those plants which are adapted to shady habitats replace the spring flora. They are like Phryma, Cryptotaenia, Tovara, Adenocaulon, Circaea, etc., along with a few evergreen plants like Pachysandra, Pyrola and Hepatica, which continue metabolism till the autumn, when they mature fruits.
(3) Taxonomic differentiation.
Since the taxonomic differentiation in plants takes place in highly intricated and very diversified mode and tempo, it is not intended here to deal with this aspect of the Arctic Tertiary floral elements on account of the space considerations.
The Arctic Tertiary floral elements show the morphological differences in varying degrees between the eastern Asiatic and the North American counterpart taxa and have been expressed by plant taxonomists at three principal levels, i.e., the ranks of subspecific status (form, variety and subspecies), species and genus. There have been a handful of literature references listing up the counterpart taxa of the Arctic Tertiary floral elements at these ranks(Gray,1859,Hara ,1952,1956,Koyama&Kawano 1964,etc.).
The cytogenetical differences between such counterpart taxa, when existing in chromosome numbers, mostly appear in polyploid series. No conspicuous correlation has been moted between the morphological differences and cytogenetical differences. For instance, in Glyceria acutiflora its subsp. acutiflora of North America is a tetraploid with 2n=40 chromosomes, hence differing from Asiatic diploid subsp. japonica with 2n=20 chromosomes. Nevertheless the morphological differences between these subspecies are very trifling. In Symplocarpus foetidus forma foetidus of North America its 2n=60 chromosomes separate the form from Asiatic forma latissimus with 2n=30 chromosomes. The morphological difference between these forms here is again very small, because the former barely differs from the latter only in its spathe which does not open during the anthesis as widely as in the latter. On the contrary, in Brachyelytrum erectum, American subsp. erectum sufficiently differs from the Asiatic subsp. japonicum in the considerably larger florets of noticeably different shape, but both subspecies show the same chromosome number of 2n=22.
At the specific level, Asiatic and North American counterpart species in a number of specific pairs have shown same chromosome numbers between them, demonstrating the fact that even these vicarious taxa have differentiated morphologically to show specific segregation the chromosomes still remain undifferentiated. For example, both Panax sching-sen of Asia and its closest ally, P. quinquefolium of North America, possess 2n=48 chromosomes, and both of Coptis japonica of Asia and its closest ally, C. asplenifolia of western North America, bear 2n=18 chromosomes.
As the examples of the generic pairs in these topics the case of Tripetaleia of Japan vs. Elliottia of the United States is often referred to. In these morphologically similar genera, which still retain certain primitive characters of the family Ericaceae, Tripetaleia can easily be separated from Elliottia in its trimerous flowers in contrast to the tetramerous ones in the latter. In Liliaceae, the genus Heloniopsis of Asia closely resembles the genus Helonias of North America, differing from the latter only in its bracted flowers, for in the latter flowers are not subtended by any conspicuous bract. In this case of Heloniopsis vs. Helonias the morphological difference between the genera is considerably smaller than in the Ericaceous case.
The area of systematic botany of the Arctic Tertiary floral elements still leaves a number of subjects for future investigations. Very little is known about the comparative chemotaxonomy of related taxa. Isn't it interesting that only the American counterpart of Symplocarpus foetidus bears such a strong odor, and hence named ‟Skunk Cabbage,” while the American Cryptotaenia canadensis subsp. canadensis lacks the fragrance of its Asian subsp. japonica? Although many species pairs have shown the common chromosome numbers between them, neither chromosomal band analyses nor comprehensive crossing experiments between counterpart taxa have hitherto been attempted. Finally, ethno- and economic botanical investigations particularly on the American counterpart taxa of Chinese and Japanese medicinal plants should help both the exploitation of new medicinal plant resources in North America and the diversification and improvement of the Oriental medicinal plant crops (Koyama 1988).
Selected References
- Fernald, M. L. 1925. Persistence of plants in unglaciated areas of boreal America. Mem. Am. Acad. Arts Sci. 15:241-342.
- Fernald, M. L. 1931. Specific segregations and identities in some floras of eastern North America and the Old World. Rhodora 33:25-63.
- Gray, A. 1846. Analogy between the floras of Japan and that of the United States. Am. J. Sci. Arts Ⅱ, 2:135-136.
- Gray, A. 1859. Diagnostic characters of phanerogamous plants with observations upon the relations of the Japanese flora to that of North America. Mem. Am. Acad. Arts Sci. N. S. 6:377-453.
- Hara, H. 1952. Contributions to the study of variations in the Japanese plants closely related to those of Europe or North America. Pt. I. J. Fac. Sci. Univ. Tokyo Ⅲ,6:29-96 ; -Pt.Ⅱ.Do.6:341-391.1956.
- Hara, H. 1962. Racial differences in widespread species, with special reference to those common to Japan and North America. Am. J. Bot. 49:647-652.
- Hu, S. Y. 1962. Phytogeographical affinities of China and the western United States. (Abstract). Am. J. Bot. 49:676.
- Hultén, E. 1937. Outlines of the history of arctic and boreal biota during the Quarternary Period. Stockholm.
- Koyama, T.& S. Kawano. 1964. Critical taxa of grasses with North American and Eastern Asiatic distribution. Can. J. Bot. 42:859-884.
- Koyama, T. 1988. Searching of ‟Chinese” medicinal plants in North America. Proc. Int. Symp. Pl. Resources 9-10. (Yunnan).
- Kryshtofovich, A. N. 1929. Evolution of the Tertiary flora in Asia. New phytologist 28:303-312.
- Li, H. L. 1952. Floristic relationships between eastern Asia and eastern North America. Trans. Am. Phil. Soc. 42:371-429.
- Raymond, M. 1951. Sedges as material for phytogeographical studies. Mém. Jard. Bot. Montréal, No.20,1-23.


『黒船が持ち帰った植物たち』コンテンツ一覧▼ 目次(青字)をクリックすると、各文をご覧いただけます
発刊に寄せて
刊行にあたって
はしがき
1.黒船による日本及びその付近での植物採集
何時,何処で,誰が?
エイサ・グレイ教授の研究
昭和天皇による黒船採集植物の御研究
Plant collecting in Japan and its adjoining territories by the Black Ships (Abstract)
2.東アジアと北アメリカの間の植物の隔離分布について
東亜と北米の間の植物の不連続分布の4型
東亜・北米間の隔離分布の由来
東亜と北米の間で隔離分布する植物の生態
隔離分布している植物の分化
Disjunct distribution of flowering plants between eastern Asia and North America (A summary account)
3.米国北太平洋探検隊採集の日本植物の図録
An annotated catalogue of plant specimens collected in Japan by the U. S. North Pacific Exploring Expeditions(1853-1855), based mainly on the set housed in The New York Botanical Garden, U.S.A.
Ⅰタイプ標本 (Type materials)
Ⅱ琉球列島 (The Ryukyus)
Ⅲ九州 (Kyushu)
Ⅳ小笠原 (The Bonin Isls.)
V下田・横浜 (Shimoda and Yokohama)
VI北海道とその周辺 (Hokkaido and its surrounding regions)
書籍詳細
-


残部僅少
[1996/11/11]小山鐵夫 著 B5判 / 上製 / 98頁(オールカラー) /
定価1,572円(本体1,429+税)/ ISBN4-900358-41-X

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
〔花の美術館〕カテゴリリンク
- 第19回 小山鐡夫追悼記念号『黒船が持ち帰った植物たち』より
- 第18回 小山鐡夫追悼記念号『植物園の話』より
- 第17回 『MAKINO 80』東京植物同好会・牧野植物同好会八十周年記念誌
- 第16回 ハマナスの語源を探る
- 第15回 金井弘夫追悼記念号『金井弘夫著作集 植物・探検・書評』より
- 第14回 『花のある風景』
- 第13回 坂嵜信之追悼記念号『日本で育つ・熱帯花木植栽事典』より
- 第12回 楽しい花観察108選『不思議な花々のなりたち』
- 第11回 川島榮生の《ヤマアジサイの世界》
- 第10回 渡辺健二『日本産ギボウシの世界』
- 第9回 『ユリノキという木』─ 魅せられた樹の博物誌
- 第8回 『北米インディアン悲詩』/ エドワード・カーティス写真集
- 第7回 江戸時代の花たち 書物に見る江戸時代の園芸文化
- 第6回 わが国初のボタニカルアーティスト 五百城文哉(いおきぶんさい)の世界
- 第5回 『ビル・ニガントゥ』 19世紀末ネパール秘蔵の本草図譜
- 第4回 植物画家 石川美枝子の作品
- 第3回 サイエンティフィック・イラストレーター 木村政司の作品
- 第2回 植物画家 中島睦子の作品
- 第1回 『PLANTAE ASIATICAE RARIORES』







営業日カレンダー
=休業日
土曜・日曜・祝日・GW休暇・夏季休暇・冬季休暇(年末年始)
- (一社)日本公園施設業協会会員
- SP・SPL表示認定企業
- ISO9001認証取得企業



アボック社は(一社)日本公園施設業協会の審査を経て「公園施設/遊具の安全に関する規準JPFA-SP-S:2024 」に準拠した安全な公園施設の設計・製造・販売・施工・点検・修繕を行う企業として認定されています。
![]()
For the happiness of the next generations